


Содержание
Перейти к:
https://doi.org/10.35754/0234-5730-2019-64-2-211-221
Перейти к:
Введение. Нейтрофилы — наиболее многочисленная субпопуляция циркулирующих в крови лейкоцитов, они составляют первую линию защиты врожденного звена иммунной системы.
Цель обзора: обобщить основные концепции фенотипической и функциональной гетерогенности нейтрофилов.
Основные сведения. В дополнение к антимикробным функциям нейтрофилов современные представления об этом пуле клеток крови отражают несколько новых стратегий захвата и уничтожения микроорганизмов: фагоцитоз и внутриклеточную деградацию, дегрануляцию и образование внеклеточных нейтрофильных ловушек после обнаружения микроорганизмов. Нейтрофилы рассматривают как фенотипически гетерогенный пул со значительной функциональной вариабельностью. В патологических условиях они могут дифференцироваться в дискретные субпопуляции с различными фенотипическими и функциональными характеристиками. Они способны входить во взаимодействие с макрофагами, природными киллерами, дендритными и мезенхимальными стволовыми клетками, B- и T-лимфоцитами или тромбоцитами, проявлять разновекторные свойства по отношению к раковым опухолям. Нейтрофилы представляют собой фенотипический гетерогенный пул клеток крови. Они обладают высокой морфологической и функциональной вариабельностью, являются модуляторами как воспаления, так и активных эффекторов иммунных реакций. Актуальна проблема поиска молекулярных маркеров, позволяющих эффективно дифференцировать фенотипы нейтрофилов, установить степень их диагностической специфичности при различных патологиях.
Андрюков Б.Г., Богданова В.Д., Ляпун И.Н. ФЕНОТИПИЧЕСКАЯ ГЕТЕРОГЕННОСТЬ НЕЙТРОФИЛОВ: НОВЫЕ АНТИМИКРОБНЫЕ Х АРАКТЕРИСТИКИ И ДИАГНОСТИЧЕСКИЕ ТЕХНОЛОГИИ. Гематология и трансфузиология. 2019;64(2):211-221. https://doi.org/10.35754/0234-5730-2019-64-2-211-221
Andryukov B.G., Bogdanova V.D., Lyapun I.N. PHENOTYPIC HETEROGENEITY OF NEUTROPHILS: NEW ANTIMICROBIC CHARACTERISTICS AND DIAGNOSTIC TECHNOLOGIES. Russian journal of hematology and transfusiology. 2019;64(2):211-221. (In Russ.) https://doi.org/10.35754/0234-5730-2019-64-2-211-221
Нейтрофилы, или полиморфнонуклеарные нейтрофилы (ПМН) (polymorphonuclear neutrophils, PMN), являются наиболее многочисленным пулом лейкоцитов и основными клеточными компонентами врожденной иммунной системы человеческого организма и обладают как противоинфекционной, так и провоспалительной функцией [1—4]. Традиционно считалось, что нейтрофилы являются относительно гомогенной популяцией терминально дифференцированных лейкоцитов, что эти клетки, в отличие от лимфоцитов и моноцитов, обладают ограниченной универсальностью и пластичностью [1]. Однако полученные в последние годы данные показали, что ПМН представляют собой фенотипический гетерогенный пул и обладают высокой морфологической и функциональной вариабельностью [2, 3], что позволяет рассматривать нейтрофилы в качестве как важных модуляторов воспаления, так и активных эффекторов иммунных реакций [3, 4].
Еще в 1920 г. высказывались сомнения насчет гомогенности этого лейкоцитарного пула, основанные на сведениях, что циркулирующие нейтрофилы могут демонстрировать значительные различия в функциях, таких как фагоцитоз, синтез белка и окислительный обмен (цит. по [2]). Однако до недавнего времени лишь ограниченное количество исследований касалось фенотипических различий ПМН, и только благодаря обнаружению высокоспецифичных нейтрофильных маркеров стало возможным разделить эту субпопуляцию клеток на подмножества [2—4].
Целью настоящего обзора является обобщить основные концепции фенотипической и функциональной гетерогенности нейтрофилов с позиции их функций и роли в развитии физиологических и патологических реакций. Особое внимание будет уделено диагностической и иммуномодулирующей стратегиям.
В нормальных условиях нейтрофилы образуются в костном мозге из морфологически неидентифицируемых гемопоэтических стволовых клеток, способных пролиферировать и дифференцироваться в белые и красные кровяные клетки и клетки — предшественники миелопоэза (гранулоцитарно-макрофагальная колониеобразующая единица (КОЕ-ГМ) гранулоци- тарная колониеобразующая единица (КОЕ-Г) [3, 5—7]. Деление и созревание этих клеток стимулируются факторами роста (интерлейкинами, гранулоцитарно- макрофагальным колониестимулирующим фактором (ГМ-КСФ) и гранулоцитарным КСФ — основными цитокинами, регулирующим гранулопоэз в норме через рецептор гранулоцитарного колониестимулирующего фактора, что приводит к формированию миелоидных пулов делящихся клеток (миелобластов, промиелоцитов и миелоцитов) и нейтрофилов, которые в процессе дифференцировки утратили способность к делению (метамиелоцитов, палочкоядерных и сегментоядерных гранулоцитов). Неделящийся пул составляет основную массу гранулоцитарного костномозгового резерва, превышающий по численности клетки, циркулирующие в периферической крови, в 30—70 раз [5, 8, 9]. Основные функции костномозгового резерва — поддерживать гомеостаз гранулоцитов («гомеостатический гранулопоэз»), циркулирующих в крови, и мобилизовать дополнительное количество клеток («экстренный гранулопоэз») при возникающей необходимости (воспаление, инфекции, рак и другие) [10, 11]. О значении гранулопоэза для организма говорит тот факт, что клетки миелоидных пулов занимают около 2/3 объема костного мозга [10].
Для поддержания гомеостаза уровня нейтрофилов других клеток в периферической крови пролиферация и дифференцировка клеток-предшественников жестко регулируется и контролируется несколькими внутренними и внешними факторами. Например, в нишах костного мозга гемопоэтические стволовые клетки сохраняются посредством взаимодействия мембранных β-интегринов с остеобластами и внеклеточным матриксом [4, 12, 13].
После созревания ПМН попадают в периферический кровоток (5х1010—1011 клеток в сутки), мигрируют в ткани, где они выполняют свои антимикробные функции. Соответственно, эквивалентное количество стареющих нейтрофилов должно быть удалено из кровообращения для сохранения гомеостаза. Нейтрофильный гомеостаз в периферической крови строго регулируется благодаря многокомпонентному механизму поддержания баланса между ростом ПМН, их дифференцировкой и элиминацией [4, 12]. Этот баланс обеспечивает выполнение этими клетками многочисленных функций с помощью фенотипической пластичности [3, 13]. Нарушение гомеостаза периферических нейтрофилов влияет на врожденный и адаптивный иммунитет, может привести к возникновению опасных для жизни инфекций, аутоиммунным или онкологическим заболеваниям.
На протяжении всей их жизни нейтрофилы по мере созревания претерпевают фенотипические переходы, которые могут быть продиктованы местным микроокружением в кровообращении и периферических тканях. Во время развития и созревания нейтрофилы проходят ряд последовательных стадий до формирования генерации зрелых нейтрофилов [12, 13]. Гранулопоэтическая популяция нейтрофилов в костном мозге делится на 3 различных пула: стволовых клеток, митотический и постмитотический. Пул стволовых клеток состоит из недифференцированных полипотентных клеток-предшественников. Митотический пул представлен миелоидными клетками-предшественниками, такими как миелобласты, промиелоциты, метамиелоциты и миелоциты. Постмитотический пул состоит из метамиелоцитов и палочкоядерных нейтрофилов. Наконец, зрелые нейтрофилы являются полностью дифференцированной клеточной генерацией, которая выходит из костного мозга и циркулирует в периферической крови.
По мере созревания и на разных стадиях диффе- ренцировки в цитоплазме нейтрофилов появляются гранулы (азурофильные, специфические и желати- назные) и секреторные пузырьки, которые содержат антимикробные пептиды и ферменты: эластазы, мие- лопероксидазы, кателицидины, дефензины и матрикс- ные металлопротеиназы, являющиеся эффективным инструментом для реализации антибактериальных стратегий ПМН [2, 10, 12].
Гранулярный аппарат нейтрофилов необходим для выполнения ими своей роли при врожденном иммунном ответе. Биологические молекулы, входящие в состав гранул, реагируют на окружающие сигналы, стимулирующие миграцию нейтрофилов к очагу инфекции и фагоцитоз вторгающихся микробов, инициируют генерацию активных форм кислорода из НАДФН-оксидазы, протеаз и антимикробных пептидов, которые, действуя совместно, создают агрессивную внутрифагосомальную среду и вызывают микробную эрадикацию [10, 13—16].
Важность образования активных форм кислорода проявляется при хронических гранулематозных заболеваниях с дефектной окислительной функцией ПМН. Для этих пациентов характерны частые рецидивирующие бактериальные и грибковые инфекции [4, 12, 14].
Являясь важными эффекторами иммунной системы, нейтрофилы ведут постоянный мониторинг организма, выявляя признаки микробных инфекций. После их обнаружения они мигрируют через эндотелий кровеносных сосудов в очаг инфекции или воспаления с помощью механизма, известного как каскад адгезии лейкоцитов (в котором принимают также участие селектины, лиганды и интегрины), и убивают вторгшиеся микроорганизмы [2, 5, 15, 16].
Миграция нейтрофилов в ткани имеет решающее значение для защиты от микробных инфекций. Важность этой функции проявляется у больных с дефектами адгезии этих клеток, что приводит к нарушению их миграции через эндотелий кровеносных сосудов. Такие больные страдают от множественных бактериальных инфекций из-за неспособности нейтрофилов к фагоцитозу [17, 18].
После выполнения своей функции в тканях нейтро- филы подвергаются апоптозу или элиминируются посредством фагоцитоза резидентными макрофагами и дендритными клетками. Запрограммированная гибель нейтрофилов — апоптоз — критичный процесс, модулирующий их количество и функции, запуская каскад противовоспалительных реакций и снижения продукции нейтрофилов [3, 17, 19—21].
В настоящее время признаны три основные проти- вомикробные стратегии нейтрофилов: фагоцитоз, дегрануляция, и формирование нейтрофильных внеклеточных ловушек (neutrophil extracellular traps, NETs) (рис. 1). NETs представляют собой защитный механизм, впервые описанный как средство улавливания и уничтожения бактерий и других патогенов [11, 22, 23]. Осуществляя такого рода внеклеточный киллинг микроорганизмов, нейтрофилы погибают через NETosis [11, 24].
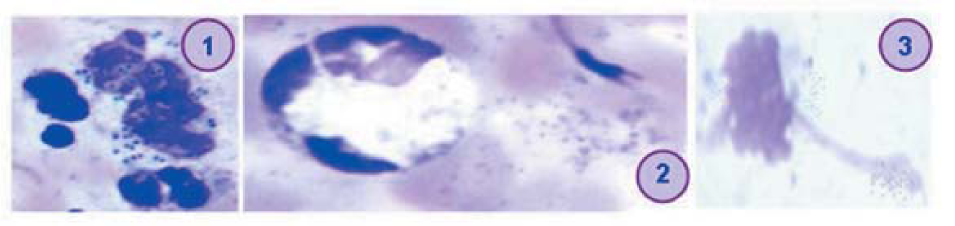
Рисунок 1. Антибактериальные стратегии нейтрофилов: 1 — фагоцитоз; 2 — дегрануляция; 3 — нейтрофильные ловушки (рисунок авторов, световая микроскопия)
Figure 1. Anfibacferial strategies of neutrophils: I — phagocytosis; 2 — degranulation; 3 — neutrophilic traps (the authors' picture, light microscopy)
Для выпуска сети активированные нейтрофилы претерпевают драматические морфологические изменения. Через минуты после активации они сплющиваются и крепко прикрепляются к субстрату. В течение следующего часа ядро теряет свои дольки, происходит деконденсация хроматина, а внутренние и внешние ядерные мембраны постепенно отделяются друг от друга. Одновременно гранулы распадаются. Через 1 ч ядерная оболочка дезагрегируется в везикулы, а нук- леоплазма и цитоплазма образуют гомогенную массу. Наконец, клетки округляются и, по-видимому, сжимаются до тех пор, пока клеточная мембрана не разрывается, а внутренняя часть клетки выбрасывается во внеклеточное пространство, образуя NETs [25].
Эти противомикробные стратегии до настоящего времени считались единственной функцией нейтро- филов. Однако недавние исследования биологии этих клеток показали, что они обладают разнообразным набором функциональных реакций, которые выходят за рамки простого уничтожения микроорганизмов [17, 18, 25].
Широкий спектр функциональных ответов нейтро- филов индуцируется активацией транскрипции и изменениями в экспрессии поверхностных молекул и их активности. Эти фенотипические изменения обычно обнаруживаются только в подгруппе нейтрофилов, что позволяет предположить наличие гетерогенных фенотипов этих клеток [3, 17, 19, 26].
Теория гетерогенности нейтрофилов зародилась более 50 лет назад, когда была предложена концепция маргинального пула нейтрофилов [4, 19, 27], либо прикрепленных к сосудам, либо находящихся полностью вне циркуляционного русла. Они могут быть мобилизованы при стимуляции [5, 28]. Эксперименты проводились с использованием радиоактивной метки нейтрофилов и помогли определить в качестве резервуаров для них селезенку, печень и костный мозг, так как требуется меньше времени для прохождения через эти органы. Вопрос о легких в качестве резервуара для нейтрофилов остается открытым [12, 26, 29]. При оценке гетерогенности нейтрофилов могут быть исследованы такие параметры, как функции клеток, маркеры клеточной поверхности, зрелость, плотность и локализация. Концепция подмножества нейтрофилов справедлива при заболевании раком, инфекционном процессе, аутоиммунитете, воспалении и репарации [27, 28].
В нормальных условиях нейтрофилы остаются в кровообращении всего несколько часов, прежде чем начинают мигрировать в ткани [30]. В течение этого времени нейтрофилы, по-видимому, меняют свой фенотип. Наблюдения каждые 4 часа за циркулирующими у здоровых мышей нейтрофилами в течение дня, показали, что эти клетки меняют свою морфологию и фенотип [21, 30, 31].
Известно много описанных фенотипов нейтрофилов, но в литературе приводится отличная друг от друга информация, поскольку в каждом опыте исследовались разные функции, использовались различные методы изучения, виды ткани, биомаркеры [32, 33].
J.M. Adrover и соавт. [22] обратили внимание на циркадные ритмы изменения функционализации нейтрофилов. Недавно вышедшие из костного мозга нейтрофилы («свежие» клетки) претерпевали несколько метаболических изменений, которые накапливались до начала миграции из кровотока в ткани. Авторы назвали эти изменения в фенотипе гранулоцитов в отсутствие воспаления старением [22].
Нейтрофилы считаются дифференцированными клетками, которые прогрессируют от незрелых форм в костном мозге до циркуляции зрелых неактивных клеток. При последующей активации во время воспалительных состояний они перемещаются из сосудов в ткани и выполняют свои функции (например фагоцитоз, производство активных форм кислорода и уничтожение бактерий) [34]. Нейтрофилы могут быстро изменять свои характеристики и манеру поведения по мере активации, созревания или изменения внешней среды [35]. Кроме того, активность нейтро- филов в ответ на различные медиаторы при остром или хроническом воспалительном процессе может продлить жизнь клетки, дать возможность произвести альтернативные эффекторные и иммунорегуляторные молекулы [34, 35].
Наконец, нейтрофилы играют совершенно полярные роли в различных врожденных иммунных реакциях, возникающих в ответ на стерильную травму, инфекцию, рак, а также в реакциях аутоиммунитета. Поэтому неудивительно, что нейтрофилы с многочисленными разновекторными функциями были идентифицированы в последние годы в разных условиях [33, 36, 37].
Такая пластичность функций нейтрофилов, отмеченная и российскими учеными [32], побудила исследователей изучить область фенотипической и функциональной гетерогенности нейтрофилов, как это было сделано для лимфоцитов [38]. Появилась возможность вычленить регуляторные, супрессорные, провоспали- тельные, воспалительные, противоопухолевые, про- опухолевые и гибридные субпопуляции нейтрофилов [39]. Помимо классического антагонистического участия в антимикробных реакциях, нейтрофилы также играют значительную роль в модуляции различных типов раковых опухолей.
В вопросах влияния на опухолевый процесс нейтрофилы в течение длительного времени были обделены вниманием исследователей по сравнению с другими иммунокомпетентными клетками [39, 40]. Одной из причин этого, по-видимому, является небольшая продолжительность жизни гранулоцитов, которая, как считалось, слишком мала (в среднем 7 ч в циркулирующей крови и еще около 5 суток в тканях), чтобы влиять на прогрессию рака [41].
Однако в публикации М. Kurashige и соавт. [41] были подтверждены результаты экспериментальных исследований Y. Sawanobori и соавт. [42] с использованием молекулярных маркеров, которые позволили пересмотреть и эти фундаментальные положения [41, 42].
Экспериментальные исследования, выполненные на животных, показали, что продолжительность жизни тканевых нейтрофилов увеличивается на несколько дней в опухолях, микроокружение которых способствует их выживанию как локально, так и системно. Установлено, что средний период продолжительности жизни циркулирующих нейтрофилов у больных раком увеличен до 5,4 дня [44, 45], что более чем в 20 раз больше, чем было обнаружено ранее [44], и расценено исследователями как следствие передачи сигналов опухолевыми цитокинами на Г-КСФ [42].
Использование современных молекулярно-генетических инструментов позволило получить новые данные об участии ПМН на всех стадиях процесса канцерогенеза: от инициации опухолевого генотипа до инвазивного роста и метастазирования злокачественной опухоли [39, 40, 46]. По результатам этих исследований установили ведущую роль нейтрофилов в канцерогенезе, связанную с их способностью изменять поведение других иммунокомпетентных клеток в развитии опухолевого процесса [46].
Изменения функций нейтрофилов и их фенотипическая гетерогенность при раке являются в настоящее время наиболее изученными и наиболее интересными метаморфозами. Эти клетки крови играют важную и противоречивую роль в развитии раковых опухолей, что отражено в ряде современных обзоров [39, 40, 43, 46-49].
Этот функциональный дуализм нейтрофилов по отношению к опухолям в течение длительного времени был предметом споров, в которых оппоненты приводили убедительные доказательства в пользу как про-, так и противоопухолевой роли этих гранулоцитов. Это привело к возникновению гипотезы о неоднородности популяции ПМН и существовании нескольких субпопуляций, функции которых при раковых опухолях могут значительно отличаться. На всех этапах развития рака нейтрофилы формируют разные клеточные фенотипы, которые выполняют разновекторные функции. Установлено, что эти гранулоциты в организме опосредуют про- (клеточный фенотип N2) и противоопухолевые реакции (фенотип N1), которые вызывают прогрессию или ингибирование опухолевых клеток [47, 48].
С одной стороны, клетки N2 способствуют росту опухоли [39, 47], ангиогенезу — новообразованию в опухоли сети кровеносных сосудов [49] и активируют метастазирование опухолевых клеток [48], индуцируя иммунодепрессию [40] и инициируя подготовку тканей к раковой колонизации — создание предметастатической ниши [50]. С другой стороны, клетки N1 проявляют противоопухолевую активность, уничтожая раковые клетки непосредственно [51, 52] либо через антителозависимую цитотоксичность, ограничивающую распространение опухоли [33, 40, 46].
Индукция этих фенотипов ПМН регулируется сигналами, исходящими от раковых или стромаль- ных клеток из микроокружения опухоли, связана как с местными, так и системными реакциями и в настоящее время является предметом пристального внимания и изучения исследователей [33, 39, 46, 49].
Мы сосредоточимся на регуляции поведения ПМН сигналами, исходящими из микроокружения опухоли, а также на диагностическом и терапевтическом потенциале гетерогенных фенотипов миелоидных клеток. Чем же обусловлен этот функциональный дуализм нейтрофилов по отношению к раковым опухолям? В последние годы установлено, что вектор про- опухолевой активности этих клеток зависит от двух условных типов подмножеств ПМН в зависимости от их плотности: высокой (HDN, от англ. high density neutrophils), обеспечивающих противоопухолевую защиту организма (соответствует фенотипу N1), и низкой (LDN, low density neutrophils) — с проопухолевой и иммунодепрессирующей активностью (соответствует фенотипу N2), а также отношением этих клеточных фенотипов [53-56].
В практике выделения нейтрофильных кластеров HDN и LDN для последующего изучения их функциональных характеристик применяются несколько протоколов, включающих в себя сбор и очистку клеток крови человека или мыши [53, 55, 56] (рис. 2). В основе всех методов — выделение гранулоцитов путем градиентного центрифугирования разведенной периферической крови (1:5) с полисахаридами соответствующей плотности (Histopaque-1077, перколл или фи- колл — 3 мл). Кровь забирается с гепаринатом лития, эритроциты перед центрифугированием лизируются. Центрифугирование проводится при 1600 g в течение 20 мин (1000 g /30 мин) [53, 57].
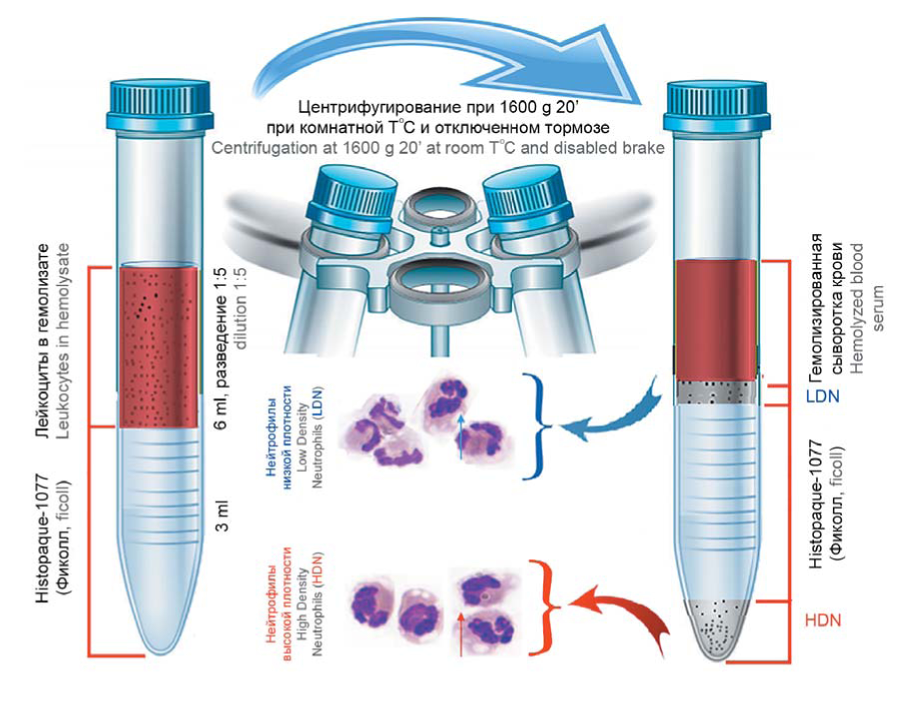
Рисунок 2. Выделение нейтрофилов высокой (HDN) и низкой (LDN) плотности из периферической крови методом градиентного центрифугирования (рисунок авторов, пояснения в тексте)
Figure 2. Isolation of neutrophils of a high (HDN) and low (LDN) density from peripheral blood by gradient centrifugation (the authors' picture, explanations in the text)
От вариаций протоколов зависит чистота выделенных клеточных фракций HDN и LDN, которые могут быть дополнительно обогащены при использовании специфических нейтрофильных антител, магнитных бус и разделительных колонок [53, 55, 56].
В последние годы интерес к субпопуляциям высокой и низкой плотности в контексте их проопухолевой активности значительно увеличился, появляется все больше данных о важной роли этих клеточных фенотипов в биологии опухолевого роста, а также механизмах регуляции раковыми клетками индукции N2-нейтрофилов [39, 53, 58, 59].
В связи с этим концепции о терминальной дифференцировке ПМН и их функциях, ограниченных антимикробной защитой и участием в воспалениях, постепенно пересматриваются [39, 47-49, 53, 59, 60]. Например, было установлено [53, 59], что у здоровых мышей более 95 % нейтрофилов в крови представлены клетками высокой плотности (N1), в то время как при возникновении злокачественной опухоли (рак молочной железы, легких, мезотелиома) у них возникала активная генерация нейтрофилов низкой плотности (N2), которые становились доминирующей циркулирующей субпопуляцией. При этом в костном мозге было достаточно много HDN, и увеличение соотношения субпопуляций в пользу LDN не было следствием простого медуллярного истощения клеток высокой плотности [53].
Аналогичная картина наблюдалась и у больных опухолями различной локализации. При этом и у мышей, и у людей доминирующая субпопуляция LDN была неоднородной и состояла из зрелых и незрелых клеток, в то время как фенотип HDN был однородным и представлен только зрелыми клетками [49, 53].
Механизмы, с помощью которых опухоли индуцируют N2-нейтропоэз, до конца не изучены, хотя в ряде исследований была показана активация клеток-пред- шественников гранулопоэза (ГМ-КСФ и Г-КСФ) и прогностическое значение отношения численности LDN- кластера гранулоцитов к лимфоцитам при нескольких типах злокачественных новообразований [51, 52].
Недавние исследования R. Kanamaru и соавт. [9] показали, что внеклеточные ловушки (NETs), высвобожденные нейтрофилами LDN-кластера, выделенными из перитонеального лаважа у больных, прооперированных по поводу рака желудка, не только не уничтожали свободные раковые клетки, но и активировали их пролиферацию после прикрепления к NETs [9].
Один из механизмов индукции образования N2-нейтрофилов, выявленный указанными исследователями, связан со стимулирующим влиянием микроокружения опухоли, точнее, с трансформирующим фактором роста-β, который индуцирует созревание в костном мозге и миграцию в ткани проопухолевого фенотипа нейтрофилов N2, изменяя локальное соотношение гранулоцитов N2:N1 в пользу первых.
Функциональная дифференциация нейтрофилов приводит к появлению различных фенотипов под влиянием конкретных опухолевых факторов, среди которых наиболее изученными являются трансформирующий фактор роста-β, Г-КСФ и интерферон-β.
Таким образом, в дополнение к классическим антимикробным стратегиям, в последние годы был охарактеризован ряд других функций нейтрофилов, что вызвало большой интерес врачей и биологов в мире. Многочисленные свидетельства того, что разные фенотипы нейтрофилов обнаружены в различных тканях и при различных патологических состояниях, позволяют предположить, что действительно существуют подмножества нейтрофилов [39, 47—49, 53, 59]. Однако отсутствие прямых экспериментальных доказательств, основанных на многократных исследованиях, не дает основания утверждать о наличии гетерогенных подгрупп нейтрофилов. Поэтому большинство практикующих врачей и гематологов считают этот клеточный пул относительно гомогенной популяцией с минимальным фенотипическим и функциональным разнообразием [48, 49, 57, 58]. Среди причин этого заблуждения тот факт, что исследования нейтрофилов все еще в значительной степени определяются их морфологией [59, 60].
Недавно выявленные с помощью градиентного центрифугирования факты наличия нейтрофилов разной плотности и их разновекторного участия в физиологических и патологических состояниях организма, возможно, вызовут смену парадигмы о функциональной гомогенности нейтрофилов [50].
Пока же приведенные в обзоре результаты исследований вызывают ряд вопросов относительно фенотипической специализации нейтрофилов. Бывают ли эти клетки запрограммированными на определенный фенотип уже до выхода из костного мозга в периферическую кровь, или высокая пластичность позволяет им трансформироваться в определенные субпопуляции уже в тканях? Возможно ли перепрограммировать уже имеющиеся фенотипы? По-прежнему остается актуальной проблема поиска надежных молекулярных маркеров, позволяющих эффективно дифференцировать фенотипы нейтрофилов, установить степень их диагностической специфичности при различных патологиях. И, наконец, обозначить перспективы определения нейтрофилов как терапевтической стратегии в иммунопатологии. В любом случае, полученные результаты выявляют потенциальные направления, на которые могут быть направлены терапевтические и диагностические стратегии усиления противоопухолевой нейтрофильной активности для раннего выявления и ограничения опухолевого роста.
1. de Oliveira S., Rosowski E.E., Huttenlocher A. Neutrophil migration in infection and wound repair: going forward in reverse. Nature Reviews Immunology. 2016; 16(6): 378. DOI: 10.1038/nri.2016.49
2. Wang J., Hossain M., Thanabalasuriar A., et al. Visualizing the function and fate of neutrophils in sterile injury and repair. Science. 2017; 358(6359): 111–6. DOI: 10.1126/science.aam9690
3. Jones H. R., Robb C. T., Perretti M., et al. The role of neutrophils in inflammation resolution. Seminars in immunology. Academic Press. 2016; 289(2): 137–45. DOI: 10.1016/j.smim.2016.03.007
4. Kaur M., Singh D. Neutrophil chemotaxis caused by chronic obstructive pulmonary disease alveolar macrophages: the role of CXCL8 and the receptors CXCR1/CXCR2. Journal of Pharmacology and Experimental Therapeutics. 2013; 347(1): 173–80. DOI: 10.1124/jpet.112.201855
5. Андрюков Б.Г., Сомова Л.М., Дробот Е.И., Матосова Е.В. Антимикробные стратегии нейтрофилов при инфекционной патологии. Клин. лаб. диагностика. 2016; 12(61): 825–33. DOI: 10.18821/0869-2084-2016-61- 12-825-833
6. Wirths S., Stefanie Bugl S., Kopp H.-G. Steady-state neutrophil homeostasis is a demand-driven process. Cell Cycle. 2013; 12(5): 709–10. DOI: 10.4161/cc.23859
7. Cain D.W., Ueda Y., Holl T.M., et al. A comparison of “steady-state” and “emergency” granulopoiesis: evidence of a single pathway for neutrophil production. The Journal of Immunology, 2009; 182 (1): 87–95.
8. Cowland J.B., Borregaard N. Granulopoiesis and granules of human neutrophils. Immunological reviews. 2016; 273(1): 11–28. DOI: 10.1111/imr.12440/
9. Kanamaru R., Ohzawa H., Miyato H., et al. Neutrophil Extracellular Traps Generated by Low Density Neutrophils Obtained from Peritoneal Lavage Fluid Mediate Tumor Cell Growth and Attachment. J Vis Exp. 2018; 138. DOI: 10.3791/58201
10. Witko-Sarsat V., Pederzoli-Ribeil M., Hirsch E., et. al. Regulating neutrophil apoptosis: new players enter the game. Trends Immunol. 2011; 32: 117–24. DOI: 10.1016/j.it.2011.01.001
11. Luo D., McGettrick H.M., Stone P.C., et al. The roles of Integrins of Human Neutrophils after their migration through endothelium into intestinal matrix. PLOS One. 2015; 10(2): e0118593. DOI: 10.1371/journal.pone.0118593
12. Steinberg B.E., Grinstein S. Unconventional roles of the NADPH oxidase: signaling, ion homeostasis, and cell death. Science’s STKE. 2007; 2007(379): pe11. DOI: 10.1126/stke.3792007pe11
13. Marini O., Costa S., Bevilacqua D., et al. Mature CD10+ and immature CD10-neutrophils present in G-CSF-treated donors display opposite effects on T cells. Blood. 2016; 04: 713206. DOI: 10.1182/blood-2016-04-713206
14. Dopico X. C., Evangelou M., Ferreira R. C., et al. Widespread seasonal gene expression reveals annual differences in human immunity and physiology. Nature communications. 2015; 6: 7000. DOI: 10.1038/ncomms8000
15. Kim M. H., Yang D., Kim M., et al. A late-lineage murine neutrophil precursor population exhibits dynamic changes during demand-adapted granulopoiesis. Scientific reports. 2017; 7: 39804. DOI: 10.1038/srep39804
16. Garlichs C.D., Eskafi S., Cicha I., et al. Delay of neutrophil apoptosis in acute coronary syndromes. Journal of leukocyte biology. 2004; 75(5): 828–35. DOI: 10.1189/jlb.0703358
17. Summers C., Singh N. R., White J. F., et al. Pulmonary retention of primed neutrophils: a novel protective host response, which is impaired in the acute respiratory distress syndrome. Thorax. 2014; thoraxjnl-2013-204742. DOI: 10.1136/ thoraxjnl-2013-204742
18. Матосова Е.В., Андрюков Б.Г. Морфофункциональная характеристика защитных механизмов нейтрофилов при бактериальных инфекциях и их вклад в патогенез провоспалительных реакций. Гематология и трансфузиология. 2017; 62(4): 223–9. DOI: 10.18821/0234-5730-2017-62-4-223-229
19. Jorgensen I., Rayamajhi M., Miao E.A. Programmed cell death as a defence against infection. Nature reviews immunology. 2017; 17(3): 151. DOI: 10.1038/ nri.2016.147
20. Jorgensen I., Lopez J.P., Laufer S.A., et al. IL-1β, IL-18, and eicosanoids promote neutrophil recruitment to pore-induced intracellular traps following pyroptosis. European journal of immunology. 2016; 46(12): 2761–6. DOI: 10.1002/ eji.201646647
21. Rodriguez F.M., Novak I.T.C. What about the neutrophil’s phenotypes? Hematol Med Oncol. 2017; 2: 1–6. DOI: 10.15761/HMO.1000130
22. Adrover J.M., Nicolás-Ávila J.A., Hidalgo A. Aging: a temporal dimension for neutrophils. Trends Immunol. 2016; 37: 334–45. DOI: 10.1016/j.it.2016.03.005
23. Brinkmann V., Zychlinsky A. Neutrophil extracellular traps: is immunity the second function of chromatin? J Cell Biol. 2012; 198(5): 773–83. DOI: 10.1083/ jcb.201203170
24. Silvestre-Roig C., Hidalgo A., Soehnlein O. Neutrophil heterogeneity: implications for homeostasis and pathogenesis. Blood. 2016 127:2173-2181 DOI: 10.1182/blood-2016-01-688887
25. Horckmans M., Ring L., Duchene J., et al. Neutrophils orchestrate postmyocardial infarction healing by polarizing macrophages towards a reparative phenotype. Eur Heart J. 2017; 38: 187–97. DOI: 10.1093/eurheartj/ehw002
26. Hellebrekers P., Vrisekoop N., Koenderman L. Neutrophil phenotypes in health and disease. Eur J Clin Invest. 2018; 48 Suppl 2: e12943. DOI: 10.1111/ eci.12943
27. Sagiv J.Y., Voels S., Granot Z. Isolation and Characterization of Low- vs HighDensity Neutrophils in Cancer. Methods Mol Biol. 2016; 1458: 179–93. DOI: 10.1007/978-1-4939-3801-8_13
28. Kuhns D.B., Priel D.A.L., Chu J., et al. Isolation and Functional Analysis of Human Neutrophils. Curr Protoc Immunol. 2015; 111: 7.23.1–16. DOI: 10.1002/0471142735.im0723s111
29. Deniset J.F., Kubes P. Neutrophil heterogeneity: Bona fide subsets or polarization states? J Leukoc Biol. 2018; 103(5): 829–38. DOI: 10.1002/JLB.3RI0917- 361R
30. Mortaz E., Alipoor S.D., Adcock I.M., et al. Update on Neutrophil Function in Severe Inflammation. Front Immunol. 2018; 9: 2171. DOI: 10.3389/ fimmu.2018.02171
31. Bekkering S., Torensma R. Another look at the life of a neutrophil. World J Hematol 2013; 2(2): 44–58. DOI: 10.5315/wjh.v2.i2.44
32. Нестерова И.В., Колесникова Н.В., Чудилова Г.А. и др. Новый взгляд на нейтрофильные гранулоциты: переосмысление старых догм. Часть 1. Инфекция и иммунитет. 2018; 7(3): 219–30. DOI: 10.15789/2220-7619- 2017-3-219-230
33. Galdiero M. R., Bonavita E., Barajon I., et al. Tumor associated macrophages and neutrophils in cancer. Immunobiology. 2013; 218(11): 1402–10. DOI: 10.1016/j.imbio.2013.06.003
34. Lai Guan Ng. Neutrophil: A mobile fertilizer J Exp Med. 2019; 216(1): 4–6. DOI: 10.1084/jem.20182059
35. Mishalian I., Granot Z., Fridlender Z.G. The diversity of circulating neutrophils in cancer. Immunobiology. 2017; 222(1): 82–8. DOI: 10.1016/j. imbio.2016.02.001
36. McCracken J.M., Allen L.A.H. Regulation of Human Neutrophil Apoptosis and Lifespan in Health and Disease. J Cell Death. 2014; 7: 15–23. DOI: 10.4137/ JCD.S11038
37. Je Lin Sieow, Sin Yee Gun, Siew Cheng Wong The Sweet Surrender: How Myeloid Cell Metabolic Plasticity Shapes the Tumor Microenvironment. Front Cell Dev Biol. 2018; 6: 168. DOI: 10.3389/fcell.2018.00168
38. Sionov R.V., Fridlender Z.G., Granot Z. The multifaceted roles neutrophils play in the tumor microenvironment. Cancer Microenviron. 2014; 8(3):125–58.
39. Coffelt S.B., Wellenstein M.D., de Visser K.E. Neutrophils in cancer: neutral no more. Nature Reviews Cancer. 2016; 16(7): 431. DOI: 10.1038/nrc.2016.52/
40. Swierczak A., Mouchemore K.A., Hamilton J.A., Anderson R.L. Neutrophils: important contributors to tumor progression and metastasis. Cancer Metastasis Rev. 2015; 34(4): 735–51. DOI: 10.1007/s10555-015-9594-9
41. Kurashige M., Kohara M., Ohshima K., et al. Origin of cancer-associated fibrobla sts and tumor-associated macrophages in humans after sex-mismatched bone marrow transplantation. Commun Biol. 2018; 1: 131.
42. Sawanobori Y., Ueha S., Kurachi M., et al. Chemokine-mediated rapid tur nover of myeloid-derived suppressor cells in tumor-bearing mice. Blood. 2008; 111(12): 5457–66. DOI: 10.1182/blood-2008-01-136895
43. Granot Z., Henke E., Comen E.A., et al. Tumor entrained neutrophils inhibit seeding in the premetastatic lung. Cancer Cell. 2011; 20(3): 300–14.
44. Shaul M.E., Levy L., Sun J., et al. Tumor-associated neutrophils display a distinct N1 profile following TGFβ modulation: A transcriptomics analysis of pro- vs. antitumor TANs. Oncoimmunology. 2016; 5(11): e1232221. DOI: 10.1080/2162402X.2016.1232221
45. Skendros P., Mitroulis I., Ritis K. Autophagy in Neutrophils: From Granulopoiesis to Neutrophil Extracellular Traps. Front Cell Dev Biol. 2018; 6: 109.
46. Rosales C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front Physiol. 2018; 9: 113. DOI: 10.3389/fphys.2018.00113
47. Uribe-Querol E., Rosales C. Neutrophils in Cancer: Two Sides of the Same Coin. J Immunol Res. 2015; 2015: 983698. DOI: 10.1155/2015/983698
48. Sag iv J. Y., Michaeli J., Assi S., et al. Phenotypic diversity and plasticity in circulating neutrophil subpopulations in cancer. Cell reports. 2015; 10(4): 562–73. DOI: 10.1016/j.celrep.2014.12.039
49. Lee W., Ko S.Y., Mohamed M.S., et al. Neutrophils facilitate ovarian cancer premetastatic niche formation in the omentum. J Exp Med. 2019; 216(1): 176–94. DOI: 10.1084/jem.20181170
50. Granot Z, Jablonska J. Distinct Functions of Neutrophil in Cancer and Its Regulation. Mediators Inflamm. 2015; 701067. DOI: 10.1155/2015/701067
51. Lopez-Lago M.A., Posner S., Thodima V.J., et al. Neutrophil chemokines secreted by tumor cells mount a lung antimetastatic response during renal cell carcinoma progression. Oncogene, 2013; 32(14): 1752–60.
52. Hong C.W. Current Understanding in Neutrophil Differentiation and Heterogeneity. Immune Netw. 2017; 17(5): 298–306.
53. Liu Y., Yue Hu, Gu F., et al. Phenotypic and clinical characterization of low density neutrophils in patients with advanced lung adenocarcinoma. Oncotarget. 2017; 8(53): 90969–78. DOI: 10.18632/ oncotarget. 18771
54. Carmona-Rivera C., Kaplan M.J. Low-density granulocytes: a distinct class of neutrophils in systemic autoimmunity. Semin. Immunopathol. 2013; 35: 455–63. DOI: 10.1007/s00281-013-0375-7
55. Wright H. L., Makki F. A., Moots R. J., et al. Low-density granulocytes: functionally distinct, immature neutrophils in rheumatoid arthritis with altered properties and defective TNF signaling. Journal of leukocyte biology. 2017; 101(2): 599–611. DOI: 10.1189/jlb.5A0116-022R.
56. Hallett M. B. The Neutrophil: Cellular Biochemistry and Physiology. CRC Press; 2017.
57. Erpenbeck L., Schön M.P. Neutrophil extracellular traps: protagonists of cancer progression? Oncogene. 2017; 36(18): 2483. DOI: 10.1038/onc.2016.406
58. Porta C., Sica A., Riboldi E. Tumor-associated myeloid cells: new understandings on their metabolic regulation and their influence in cancer immunotherapy. FEBS J. 2018; 285(4): 717–33. DOI: 10.1111/febs.14288
59. Khadge S., Sharp J.G., McGuire T.R., et al. Immune regulation and anticancer activity by lipid inflammatory mediators. Int Immunopharmacol. 2018; 65: 580–92. DOI: 10.1016/ j. intimp.2018.10.026
Андрюков Борис Георгиевич, заслуженный врач РФ, доктор медицинских наук, ведущий научный сотрудник лаборатории молекулярной микробиологии
профессор департамента фундаментальных наук
тел.: +7(924)230-46-47; 690087, г. Владивосток, ул. Сельская, д. 1.
Андрюков Б.Г., Богданова В.Д., Ляпун И.Н. ФЕНОТИПИЧЕСКАЯ ГЕТЕРОГЕННОСТЬ НЕЙТРОФИЛОВ: НОВЫЕ АНТИМИКРОБНЫЕ Х АРАКТЕРИСТИКИ И ДИАГНОСТИЧЕСКИЕ ТЕХНОЛОГИИ. Гематология и трансфузиология. 2019;64(2):211-221. https://doi.org/10.35754/0234-5730-2019-64-2-211-221
Andryukov B.G., Bogdanova V.D., Lyapun I.N. PHENOTYPIC HETEROGENEITY OF NEUTROPHILS: NEW ANTIMICROBIC CHARACTERISTICS AND DIAGNOSTIC TECHNOLOGIES. Russian journal of hematology and transfusiology. 2019;64(2):211-221. (In Russ.) https://doi.org/10.35754/0234-5730-2019-64-2-211-221
![]()
125167, Москва, Новый Зыковский проезд, 4
ФГБУ «НМИЦ гематологии» Минздрава России
тел.: 8-926-816-3887
e-mail: o.levchenko@htjournal.ru






































